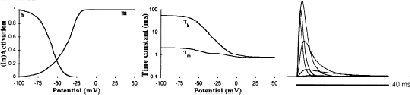Related Documentation:
The presence of a conductance like an A current (KA) in the Purkinje cell was shown by [6]. We derived our equations for KA from the original reports on A currents [2, 3] modified to fit the data in the two reports on Purkinje cell A currents that were available at that time. The whole cell voltage-clamp study of cultured Purkinje cells by [5] (their Fig. 9) provided data about the activation and inactivation time constants and the steady-state inactivation, and a single-electrode voltage-clamp study in slice by [7] supplied steady-state activation data.
Recently [9] published a more complete report on a single-electrode voltage-clamp study of the Purkinje cell. The average threshold for activation they report is lower than the value used in our model, but there seems to be a large natural variability in the activation curves (compare their Fig. 1 with their Table 1). The steady-state inactivation curve of [9] is similar to ours and to the kinetics of A currents in other systems [2, 8] but they report a much slower inactivation time constant than [5].
One point of contention in the literature about the distribution of ionic channels involves the presence or absence of an A current in the distal dendrites [1, 6]. One argument for a more extensive distribution is that a depolarizing bias current, which would inactivate A currents, also causes the dendrite to fire spikes more easily. However, a similar result would be expected if the depolarization also activated a plateau current, as shown in Fig. 11. Moreover, in contrast to the expected 4-aminopyridine (4-AP) sensitivity of A currents [8], the outward current described by [6] was not blocked by 4-AP. In addition, voltage-clamp data [9] do not support a distal dendritic location of the A current. Patch-clamp studies of cultured Purkinje cells [4] have shown the A current to be present in both somatic and dendritic membrane, but it is likely that patches were obtained from smooth dendrites only. In the current model an A current was present in the soma and main dendrite. However, dendritic spiking was more influenced by the K2 Ca2+ -activated K+ current, which allowed a finer control of dendritic excitability than the A current, which inactivated quickly during long current injections.
[1] CY Chan, J Hounsgaard, and C Nicholson. Effects of electric fields on transmembrane potential and excitability of turtle cerebellar Purkinje cells in vitro. Journal of Physiology (Lond.), 401:751–771, 1988.
[2] JA Connor and CF Stevens. Voltage clamp studies of a transient outward membrane current in gastropod neural somata. Journal of Physiology (Lond.), 213:21–30, 1971.
[3] E De Schutter. Alternative equations for the molluscan ion currents described by connor and stevens. Brain Research, 382:134–138, 1986.
[4] DL Gruol, T Jacquin, and AJ Yool. Single-channel K+ currents recorded from the somatic and dendritic regions of cerebellar purkinje neurons in culture. Journal of Neuroscience, 11:1002–1015, 1991.
[5] T Hirano and S Hagiwara. Kinetics and distribution of voltage-gated Ca, Na, and K channels on the somata of rat cerebellar Purkinje cells. Pfluegers Archiv, 413:463–469, 1989.
[6] J Hounsgaard and J Midtgaard. Intrinsic determinants of firing patterns in Purkinje cells of the turtle cerebellum in vitro. Journal of Physiology (Lond.), 402:731–749, 1988.
[7] SJ Li, Y Wang, HK Strahlendorf, and JC Strahlendorf. A transient voltage-dependent outward current recorded from rat cerebellar purkinje cells under voltage clamp. Society for Neuroscience Abstracts, 16:507, 1990.
[8] MA Rogawski. The A-current: How ubiquitous a feature of excitable cells is it? Trends in Neurosciences, 8:214–219, 1985.
[9] Y Wang, JC Strahlendorf, and HK Strahlendorf. A transient voltage-dependent outward potassium current in mammalian cerebellar purkinje cells. Brain Research, 567:153–158, 1991.