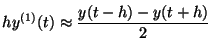In the one dimensional cable equation the membrane potential is dependent on time as well as space. Both these axes can be discretized independently. We first assume that the cell is iso-potential (same membrane potential at all places) which eliminates the spatial dependence (if you omit the Hodgkin-Huxley conductances, this also leads to integrate-and-fire models).
Since the cell is assumed to be iso-potential, the cable equation reduces to a simple exponential decay function. The question remains how to fill in the Hodgkin-Huxley current while maintaining the second-order accuracy of the trapezoidal rule. The following trickery is used for this:
Subtract and regroup:
Or if you rewrite:
This equation has a local truncation error of third order and can be
used to fill in the membrane potential in the Hodgkin-Huxley equations
and vice versa in the following way: we assume that ![]() is the
unknown gate value we are searching, while
is the
unknown gate value we are searching, while ![]() is the previous
gate value and
is the previous
gate value and
![]() is the rate of change for the gate value
that can be calculated given the membrane potential at time
is the rate of change for the gate value
that can be calculated given the membrane potential at time ![]() .
Under the assumptions that
.
Under the assumptions that ![]() and the membrane potential are
second-order correct, the result of this calculation - the new values
for the gates - will be second-order correct too. Then the same
method can be used to compute the membrane potential at time point
and the membrane potential are
second-order correct, the result of this calculation - the new values
for the gates - will be second-order correct too. Then the same
method can be used to compute the membrane potential at time point
![]() given the membrane potential at time point
given the membrane potential at time point ![]() and the
gate values at time point
and the
gate values at time point ![]() .
.
It is important to see that the second-order accuracy is maintained only when the membrane potential and the Hodgkin-Huxley equations are computed at different time points.
| NOTE: If concentration pools are represented in a model of a neuron, they should be evaluated at the same time steps as the membrane potential. |
![\includegraphics[scale=0.6]{figures/mstep.eps}](img49.png) |